
Bienvenue sur NéoSVT. Ce chapitre s’inscrit dans l’unité de production de la matière organique et des flux d’énergie. Nous allons explorer comment les plantes chlorophylliennes interagissent avec le sol pour puiser les ressources minérales nécessaires à leur métabolisme.
Introduction
La plante utilise l’eau, les sels minéraux et le CO2 pour synthétiser sa propre matière organique grâce à l’énergie lumineuse et la chlorophylle (Photosynthèse).
- Par quels mécanismes absorbent-elles ces éléments ?
- Comment s’effectuent les échanges gazeux et l’utilisation de la lumière ?
I- Mise en évidence des échanges d’eau chez les végétaux chlorophylliens
1- Nécessité d’eau et des sels minéraux
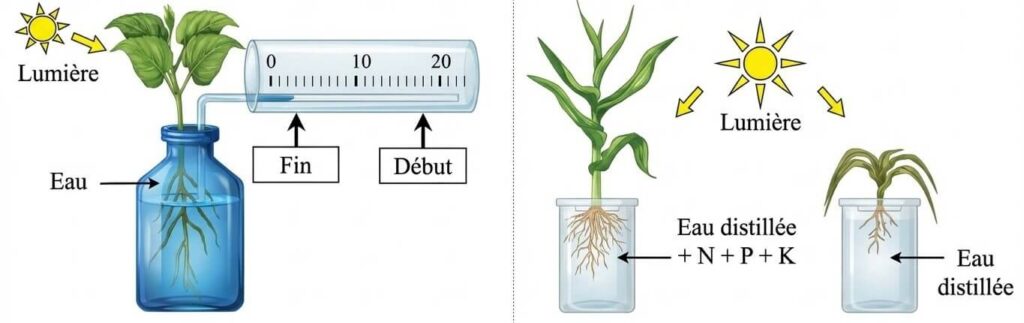
L’analyse expérimentale montre que les végétaux absorbent l’eau en permanence par leurs racines. Une croissance optimale nécessite un mélange équilibré de sels minéraux (N, P, K). Sans ces éléments, le développement est limité.
2- Observations macroscopiques : Expérience de la pomme de terre
En plaçant des morceaux de pomme de terre (30 mm) dans différentes concentrations de saccharose, nous observons les résultats suivants :
| Concentration (mol/L) | 0 (Eau distillée) | 0.1 | 0.2 | 0.3 | 0.4 | 0.5 |
|---|---|---|---|---|---|---|
| Longueur finale (mm) | 32 | 31 | 30 | 29 | 28 | 27 |
| État de la cellule | Turgescence | Turgescence | Équilibre | Plasmolyse | Plasmolyse | Plasmolyse |
Conclusion : Les tissus végétaux absorbent l’eau en milieu hypotonique (< 0.2 mol/L) et en perdent en milieu hypertonique (> 0.2 mol/L).
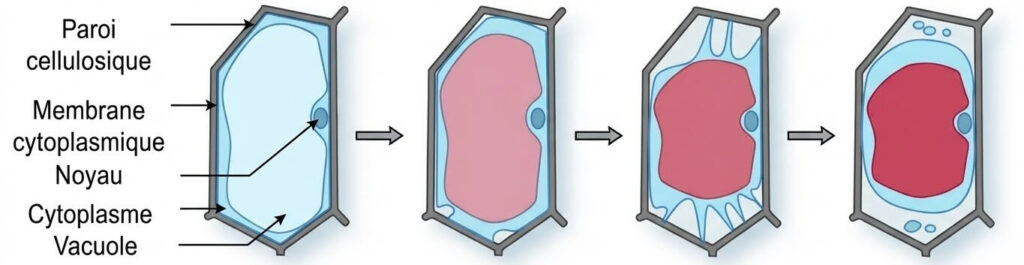
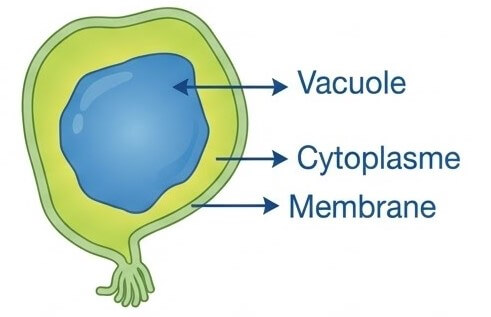
3- Rappel : Structure de la cellule végétale
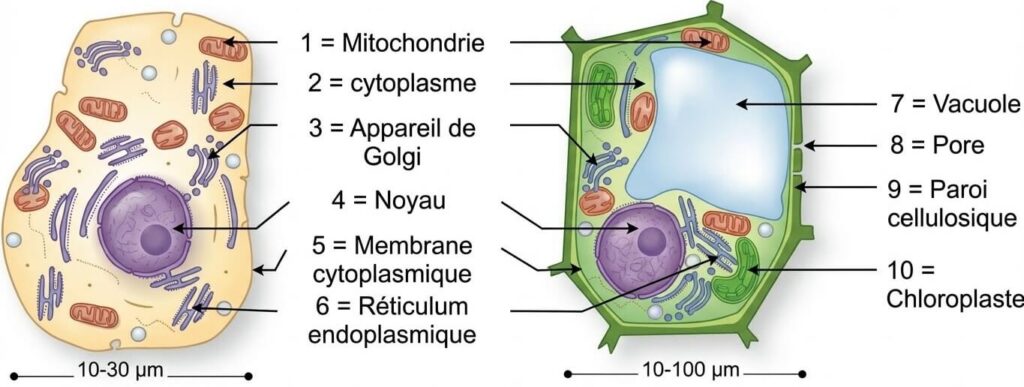
La cellule végétale se distingue de la cellule animale par sa forme polyédrique, sa paroi pectocellulosique rigide, la présence de chloroplastes et de vacuoles centrales de grande taille.
4- Observations microscopiques : Cellules d’oignon
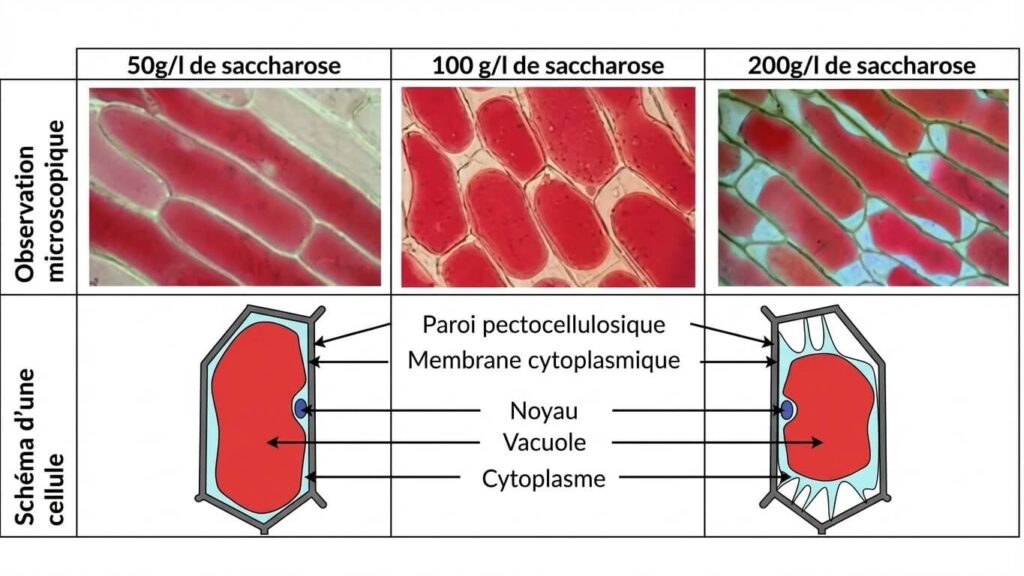
- Milieu hypotonique (50g/l) : La cellule est en état de turgescence. La vacuole est gonflée d’eau et presse le cytoplasme contre la paroi.
- Milieu isotonique (100g/l) : La cellule garde un aspect normal.
- Milieu hypertonique (200g/l) : La cellule est plasmolysée. La vacuole se rétracte suite à une perte d’eau.
II- Modèle physique pour expliquer les échanges d’eau
1- Notion d’osmose : Expérience de Dutrochet
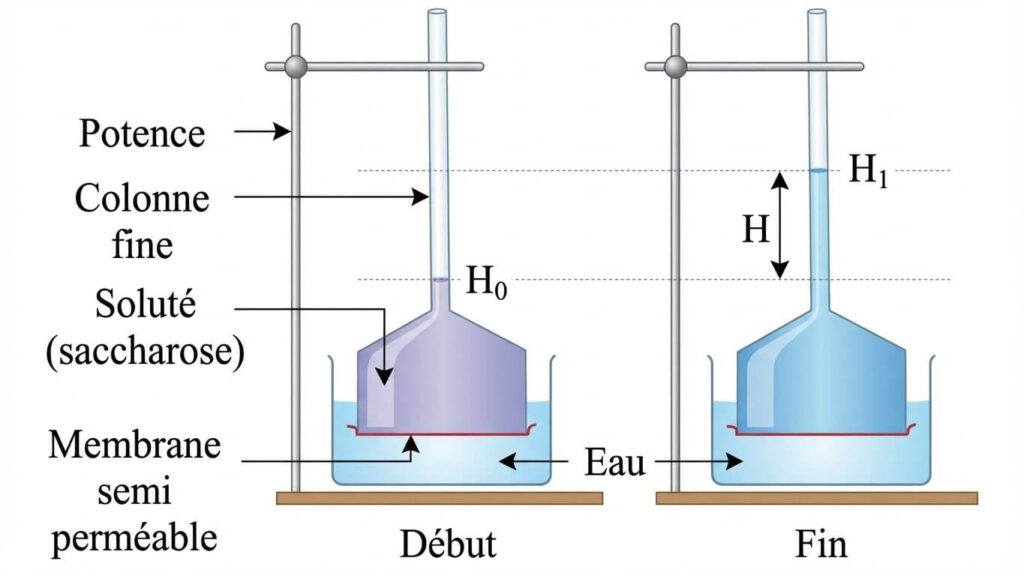
L’osmomètre montre que l’eau traverse une membrane semi-perméable du milieu le moins concentré (hypotonique) vers le plus concentré (hypertonique). C’est le phénomène d’osmose provoqué par la pression osmotique.
2- Calcul de la pression osmotique
π = i . R . T . C
P : Pression osmotique(atm) | i : Nombre de particules produites par la dissociation d’une unité de soluté. | R : Constante des gaz parfaits (0.082) | T : Température absolue (K) | C : Concentration molaire (mol/l)
✍️ Application :
Énoncé : On dissout 540 mg de glucose (C6H12O6) dans 30 ml d’eau à 25°C.
Données : M(O)=16; M(C)=12; M(H)=1 g/mol ; R = 0,082
- Calculer la masse molaire du glucose.
- Calculer la concentration massique (Cm) et molaire (C).
- Calculer la pression osmotique (π) de cette solution.
✅La Correction :
1. Masse molaire (M) : (6 × 12) + (12 × 1) + (6 × 16) = 180 g/mol
2. Concentrations :
• Cm : m / V = 0.54g / 0.03L = 18 g/l
• C : Cm / M = 18 / 180 = 0.1 mol/l
3. Pression osmotique (π) :
π = 1 × 0.082 × (25 + 273) × 0.1 = 2.4436 atm
2- Calcul de la concentration molaire C et et la concentration massique Cm à l’isotonie .
Pour calculer la concentration d’une solution 1 isotonique à une solution 2 on utilise la formule suivante : π1 = π2
Donc : i1.R.T.C1 = i2.R.T.C2
En simplifiant R et T, on obtient l’égalité fondamentale :
i1.C1 = i2.C2
Une fois que vous avez C2, vous pouvez trouver la concentration molaire C1 :
Vous pouvez aussi trouver la concentration massique Cm (en g/L) en utilisant la masse molaire (M1) du soluté :
vous pouvez voir cette exercice d’application dans notre chaine youtube cliquez ici
III. Les mécanismes d’absorption des sels minéraux
L’absorption s’effectue sous forme d’ions selon deux mécanismes : la diffusion et le transport actif.
1. La diffusion (Transport passif)
Passage des ions du milieu le plus concentré vers le milieu le moins concentré, sans dépense d’énergie.
A. La diffusion simple
Passage libre à travers la bicouche phospholipidique, proportionnel au gradient de concentration.
B. La diffusion facilitée
Utilise des protéines spécifiques (transporteurs). Ce mécanisme est rapide mais saturable.
| Caractéristique | Diffusion Simple | Diffusion Facilitée |
|---|---|---|
| Sens du transport | Selon le gradient | Selon le gradient |
| Protéines porteuses | Non requises | Requises (Transporteurs) |
| Énergie (ATP) | Non (Passif) | Non (Passif) |
| Vitesse / Saturation | Lente / Non saturable | Rapide / Saturable |
C. La diffusion orientée
Mise en évidence par le rouge neutre : le colorant pénètre mais ne ressort pas. La diffusion ne se fait que dans un seul sens.
2. Le transport actif : Un mécanisme dépendant de la vie cellulaire
Étude de cas : L’analyse des concentrations ioniques chez l’algue marine Valonia ventricosa révèle un mode de transport opposé à la diffusion.
| Ions | Eau de mer (Extérieur) | Suc vacuolaire (Intérieur) | Sens du gradient |
|---|---|---|---|
| Na+ | Élevée | Faible | Extérieur → Intérieur |
| K+ | Faible | Élevée | Intérieur → Extérieur |
🔍 Analyse et Interprétation :
1. Déséquilibre ionique : Les mesures révèlent une répartition asymétrique. Le sodium est plus abondant à l’extérieur, alors que le potassium est accumulé à l’intérieur.
2. Opposition à la diffusion : Le maintien permanent de ce gradient prouve que la cellule s’oppose à l’équilibre spontané par un mécanisme actif.
💡 Conclusion : Le Transport Actif
- À contre-courant : Se fait du milieu le moins concentré vers le plus concentré.
- Consommation d’ATP : Nécessite de l’énergie cellulaire.
- Vitalité : Bloqué par le cyanure (inhibiteur d’énergie).
Synthèse Bilan : La membrane plasmique puise activement les éléments nécessaires grâce à ses protéines membranaires (pompes).
3. Caractéristiques de perméabilité de la membrane cytoplasmique
Le comportement cellulaire en milieu hypertonique révèle les propriétés suivantes :
- 🚀 Perméabilité sélective : Cas du saccharose. Membrane imperméable (pas de déplasmolyse).
- ⚖️ Perméabilité différentielle : Membrane plus perméable au glucose qu’au NaCl.
- ⚡ Perméabilité élevée : Cas de l’urée et du glycérol (déplasmolyse immédiate).
IV- Structures responsables de l’absorption
1- Les poils absorbants
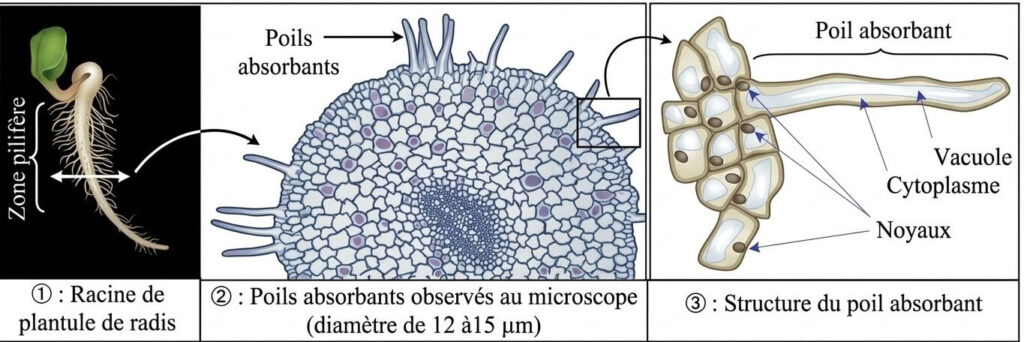
L’absorption se fait au niveau de la zone pilifère de la racine où il y a les poils absorbants. Ils augmentent la surface d’échange et possèdent une forte pression osmotique.
2- Rôle de la transpiration
L’évaporation au niveau des feuilles crée une force d’aspiration qui tire la sève brute vers les feuilles.
V- Structures cellulaires intervenant dans l’absorption
1- La paroi pectocellulosique
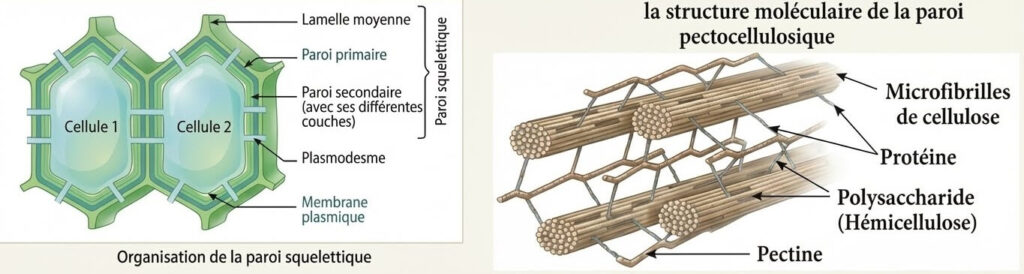
Rigide et perméable grâce aux plasmodesmes qui permettent la communication entre les cellules.
2- La membrane plasmique : Modèle de la mosaïque fluide
Modèle de Singer et Nicholson (1972) : Une bicouche phospholipidique dynamique avec des protéines mobiles assurant le contrôle des flux.
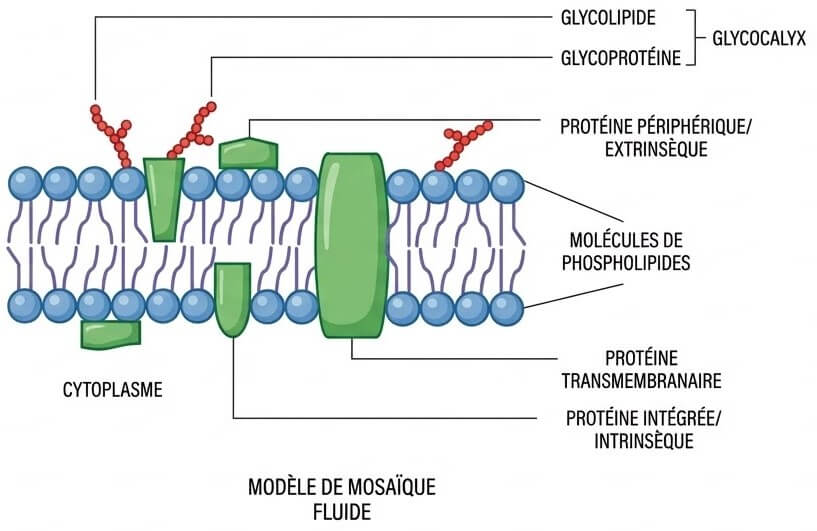
VI- Bilan : Le contrôle de l’entrée de l’eau et des sels
Pour atteindre la vacuole, les molécules franchissent différentes barrières :
Étape 1 La Paroi : Passage libre
Totalement perméable, elle laisse circuler librement l’eau et les solutés vers la membrane.
Étape 2 La Bicouche Lipidique : Filtre à eau
Responsable de l’Osmose et de la diffusion simple.
Étape 3 Les Protéines : Contrôle sélectif
Permettent la diffusion facilitée et le Transport actif (via ATP).
| Cible du passage | Structure responsable | Mécanisme lié |
|---|---|---|
| Eau et Sels | Paroi | Diffusion libre |
| Molécules d’Eau | Bicouche lipidique | Osmose |
| Ions et Nutriments | Protéines membranaires | Transport Actif |
💡 Conclusion : L’absorption est un travail coordonné où la Membrane Plasmique sélectionne ce dont la plante a besoin grâce à sa structure en Mosaïque Fluide.